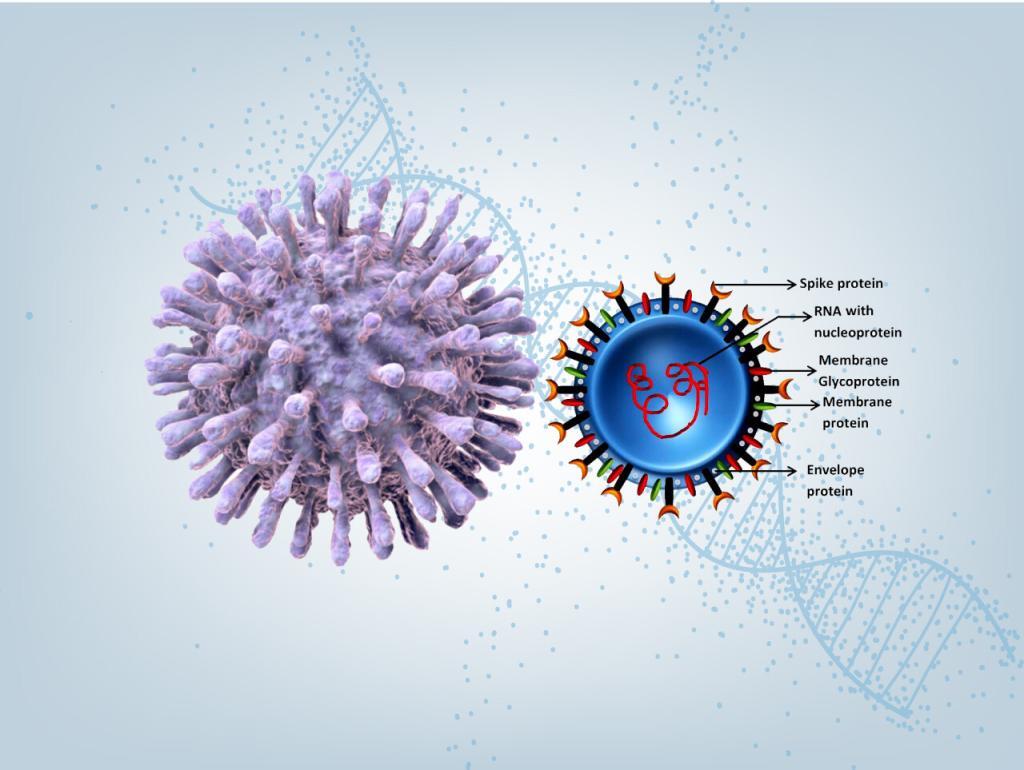
2019 Novel Coronavirus SARS-CoV-2 is a virus identified as the cause of an outbreak of respiratory illness Covid-19 first detected in Wuhan, China.
To help expedite Covid-19 research, LifeTein synthesized a 69 amino acid spike glycoprotein with one disulfide bond in 6 days. This effort is a partnership with a biotech company for drug development.
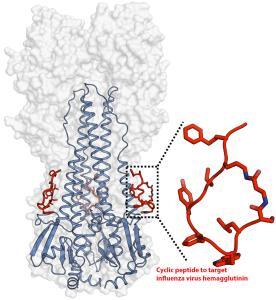
Cyclic peptides as broad-spectrum antiviral agents
Antiviral drugs and vaccines are the most powerful tools to combat viral diseases. Most drugs and vaccines only target a single virus. However, the broad-spectrum antivirals can be used for rapid management of new or drug-resistant viral strains. Cyclized peptides and peptide analogs are excellent examples of broad-spectrum antivirals.
An artificial peptide molecule was found to neutralize a broad range of group 1 influenza A viruses, including H5N1. The peptide design was based on complementarity determining region (CDR) loops have been reported for other viral targets. The optimized peptides bind to the highly conserved stem epitope and block the low pH-induced conformational rearrangements associated with membrane fusion.
These peptidic compounds and their advantageous biological properties should accelerate development of novel small molecule and peptide-based therapeutics against influenza virus.
The linear peptide is Suc-SQLRSLEYFEWLSQ-NH2. Three cyclization strategies were used: head to tail, side chain to side chain and side chain to tail. An ornithine (Orn) side chain was fused with the carboxyl terminus of β-alanine for lactam formation.
Check here for more details: Potent peptidic fusion inhibitors of influenza virus, Science 28 Sep 2017, DOI: 10.1126/science.aan0516
Lately, more broad-spectrum antiviral agents were found to target viruses. It was found that 55 compounds can target eight different RNA and DNA viruses. Dalbavancin is a novel lipo-glycopeptide antibiotic. The lipoglycopeptide disrupts bacterial cell wall formation by binding to
the terminal d-alanyl-d-alanine peptidoglycan sequence in Gram-positive bacteria in a linear, concentration-dependent manner. The dalbavancin has effects on echovirus 1, ezetimibe against HIV1 and Zika virus.
More details: https://www.ncbi.nlm.nih.gov/pubmed/29698664
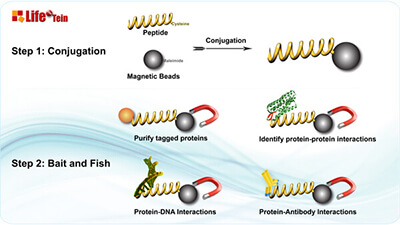
Protein Purification Using Magnetic Beads: Top Tips for Success
Magnetic bead-based protein purification offers a powerful solution for various applications like high-throughput microscale purification, pull-down/CoIP experiments, and protein-protein or protein-DNA interaction studies. Here’s why magnetic beads are the top choice: they can be coated with specific affinity ligands for antigens, antibodies, proteins, or nucleic acids. Moreover, magnetic beads are non-porous and have a defined diameter, eliminating hidden surfaces where molecules can stick, leading to reduced background, simplified purification, and streamlined washing steps. Compared to traditional bead separation methods involving agarose, sepharose, or silica beads, magnetic bead separation stands out as the quickest, cleanest, and most efficient technique.
If you’re new to working with magnetic beads, here are some essential tips to ensure success:
Thorough Resuspension: Ensure uniformity across aliquots by thoroughly resuspending your magnetic beads. These nano-superparamagnetic beads are covalently coated with highly functional groups, providing increased binding capacity and better dispersion. Since magnetic beads are composed of iron oxide and can settle over time, it’s crucial to vortex and resuspend them thoroughly before use to redisperse the beads.
Enhanced Washing: Minimize non-specific binding by increasing the number of washing steps. Whether you’re using ethanol or the recommended wash buffer, make sure to use an adequate volume of wash solution to cover the bead pellet.
Understanding Functional Groups: Different beads are covalently coated with various functional groups like maleimide, primary amine, NHS, carboxylic acid, purified streptavidin, protein A, reduced glutathione, nickel-charged nitrilotriacetic acid, or groups for DNA/RNA purification. These coatings, along with buffer conditions, affect bead properties. Understanding these specifics is essential for proper bead handling.
Efficient Bead Capture: Magnetic beads typically form a pellet attracted to the magnet within a minute. Extend the attraction time to ensure efficient bead capture.
Gentle Supernatant Removal: When removing the wash solution or supernatant, angle the pipette tip to avoid disturbing the magnetic bead pellet. Ensure that the tip doesn’t come into contact with the pellet.
By following these tips, you can make the most of magnetic bead-based protein purification, improving the efficiency and reliability of your experiments.
Giantin, a novel conserved Golgi membrane protein, is a disulfide-linked homodimer. It was found that BFA-induced Golgi disorganization is associated with the monomerization of giantin.
The pull-down experiment was performed. The control peptide biotin-GHGTGSTGSGSMLRTLLRRRL synthesized by LifeTein was incubated with lysate and Dynabeads, as well as the lysate incubated with Dynabeads only served as a control. Dynabeads carrying MGAT1 peptide were able to pull-down giantin from the lysate of HeLa cells, however, giantin was not detected in the pull-down fraction from the lysate exposed to the Dynabeads or in combination with control peptide. It is logical to hypothesize that the MGAT1 binding domain of giantin lies within its N-terminal non-coiled-coil area.
The Dynabeads function similarly to LifeTein magnetic beads: https://www.lifetein.com/peptide-product/amineactivated-peptide-conjugation-magnetic-beads-p-3647.html
LifeTein provides the fastest turnaround time and most reliable quality in the industry. Peptides are made in New Jersey, USA. Projects move from conception to bench in only 3–5 days so you can deal with your research deadlines.
Introducing LifeTein‘s faster microwave peptide synthesis technology! LifeTein’s new platform is designed for maximized speed and efficiency. Unparalleled peptide quality, greater flexibility, and improved reliability make LifeTein the vendor of choice for all your peptide synthesis needs.
Aβ-(1–42) was dissolved to 1 mM in 100% hexafluoroisopropanol, hexafluoroisopropanol was removed under vacuum, and the peptide was stored at −20 °C. For the aggregation protocols, the peptide was first resuspended in dry Me2SO (DMSO) to 5 mM. For oligomeric conditions, F-12 (without phenol red) culture media was added to bring the peptide to a final concentration of 100 μM, and the peptide was incubated at 4 °C for 24 h. For fibrillar conditions, 10 mM HCl was added to bring the peptide to a final concentration of 100 μM, and the peptide was incubated for 24 h at 37 °C.
ADDLS, amyloid-derived diffusible ligands.
Preparing human islet amyloid polypeptide (hIAPP), also known as amylin, can be challenging due to its hydrophobic amino acid residues.
Here’s an improved method for dissolving lyophilized hIAPP:
Begin by dissolving lyophilized hIAPP in 80% (v/v) HFIP containing 10 mM HCl. This step ensures complete dissolution. The CD spectrum indicates the presence of a stable alpha-helical conformation, which remains so for several days.
Next, remove the HFIP by lyophilization, leaving behind lyophilized hIAPP.
Re-dissolve the lyophilized hIAPP in 10 mM HCl, and eliminate any insoluble components by ultracentrifugation.
The resulting hIAPP solution in 10 mM HCl is ready for immediate use in experiments.
To initiate the formation of hIAPP fibrils, introduce the stock solution into the reaction buffer. Conditions for fibril formation were optimized under two pH conditions:
Low pH: Utilize 25 uM hIAPP in 10 mM HCl, with varying concentrations of HFIP.
Neutral pH: Employ 25 uM hIAPP in a 50 mM sodium phosphate buffer at pH 7.0, with varying concentrations of HFIP.
Incubate these samples at 25 °C for several hours.
Reference: JOURNAL OF BIOLOGICAL CHEMISTRY 23965, JULY 8, 2011 VOLUME 286 NUMBER 27
Aducanumab is a human monoclonal antibody that has been studied for the treatment of Alzheimer’s disease.
The Nobel Prize in Physiology or Medicine 2019 was awarded jointly to William G. Kaelin Jr., Sir Peter J. Ratcliffe, and Gregg L. Semenza. LifeTein made one of the key peptides: HIF1a-P564: DLDLEMLAPYIPMDDDFQLR.
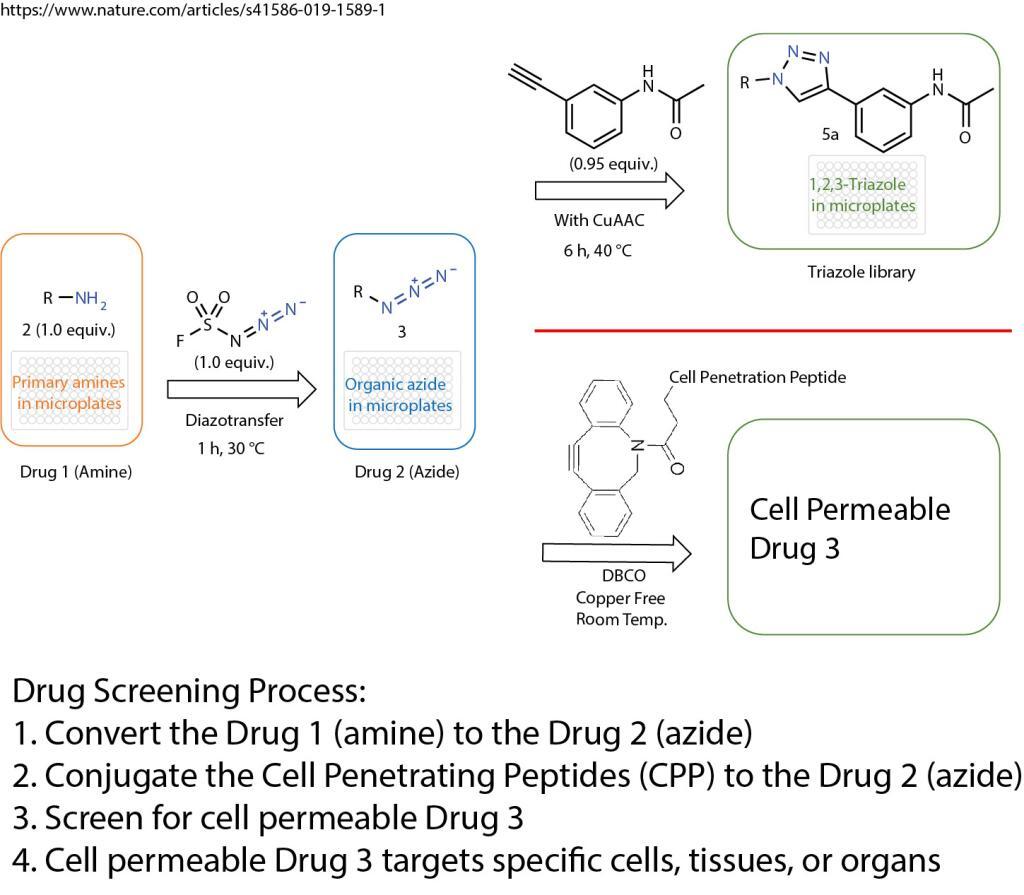
A click chemistry was reported about the formation of azides from primary amines. This powerful tool enables the reaction of just one equivalent of a simple diazotizing species, and fluorosulfuryl azide (FSO2N3), for the preparation of over 1,200 azides on 96-well plates in a safe and practical manner. This method greatly expands the number of accessible azides and 1,2,3-triazoles because the primary amine is one of the most abundant functional groups in small compounds, proteins and antibodies.
Formation of Azides From Primary Amines
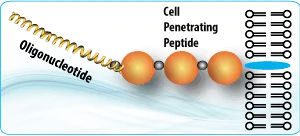
The method opens the door for numerous applications in drug screening and discovery. The cell penetration peptides can be easily introduced to conjugate with any azide containing drugs, compounds, antibodies, or proteins.
The cell penetration peptides (CPPs) are capable of delivering biologically active cargo to the cell interior. The desired therapeutic cargo could be attached to a CPP using the copper free click chemistry and then delivered to an intracellular target, thereby overcoming the entry restrictions set by the plasma membrane.
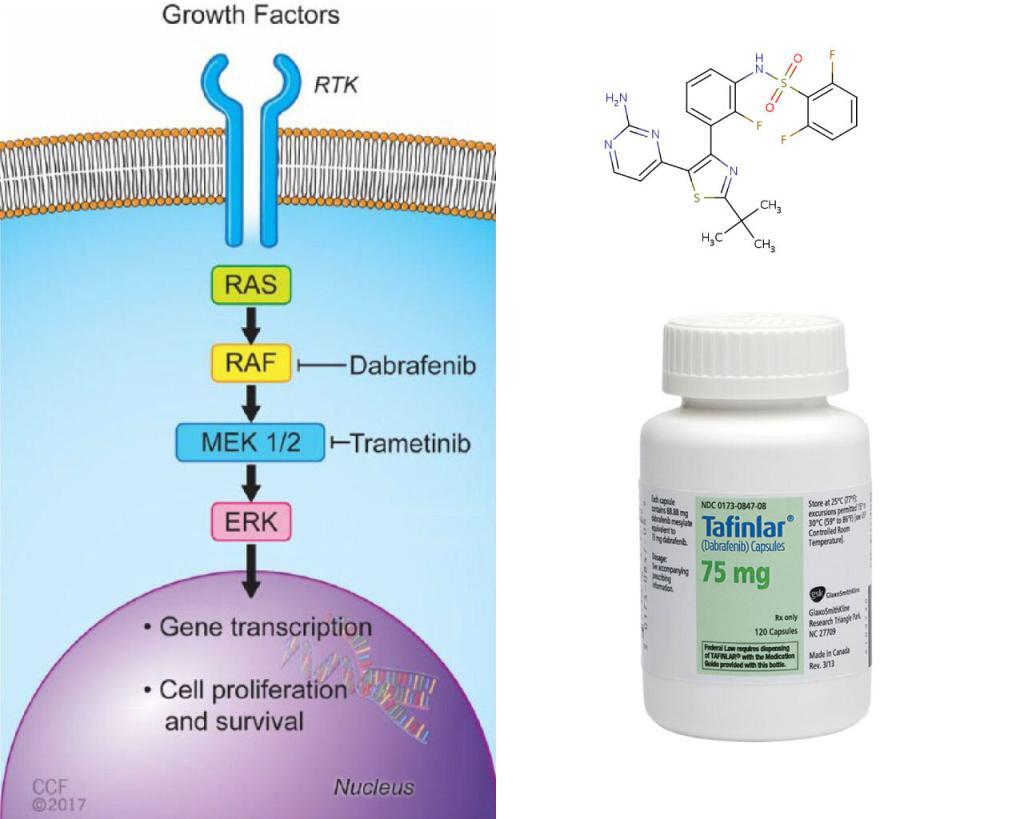
BRAF is an RAF kinase. It is a core component of the RAS/RAF/MEK/ERK signaling cascade, known as mitogen-activated protein kinase (MAPK) pathway. It is one of the major effectors of oncogene RAS, and is often mutated in human cancer cells.
LifeTein’s Braftide & Cancer Therapy
Two FDA approved drugs, Dabrafenib, and vemurafenib, effectively inhibit the most common BRAF variant V600E, a monomeric BRAF. But, the non-V600E BRAF mutations are intrinsically resistant to these drugs. These drugs may also paradoxically stimulate the pathway when the tumor cells contain wild-type BRAF and oncogenic RAS, causing secondary malignancies. The researchers tried to tackle the dimeric BRAF. The dimeric BRAF, such as the wild type and G469A, a most prevalent non-V600E variant in lung cancer cells, hinges on dimer interface (DIF), a 20aa span near the tail end of the alpha-C helix of BRAF. The researchers designed Braftide using computational modeling, aiming to block the dimerization. They tested the functionality in vitro, in HEK263 cells and colon cancer cell lines.
LifeTein synthesized Braftide (TRHVNILLFM), Null-Braftide (THHVNILLFM), Cy3-Braftide (TRHVNILLFM-Cy3), TAT-Braftide (GRKKRRQRRRPQ-PEG-TRHVNILLFM), and TAT (GRKKRRQRRRPQ). We reviewed here some of the assays that helped support Braftide as an allosteric inhibitor of BRAF dimer and down-regulator of MAPK signaling pathway for cancer therapy.
1) Cell-free in vitro assay: dose-response curve. First of all, the researchers show that Braftide has a sub-micromolar IC50 for dimeric BRAF. Full-length dimeric BRAF-WT and BRAF-G469A (from HEK293F cells) were used for dose-response curves, and the BRAF activity was probed by pMEK production.
2) Cell-free in vitro assay: Saturation binding assay. The researchers used Cy3-labeled Braftide (Cy3-Braftide) to characterize (KD) the binding of Braftide with dimeric BRAF-WT using fluorescence quantification.
3) Cell-free in vitro assay: Immunoprecipitation (IP). The purpose of IP was to show Braftide disrupted the BRAF dimerization. Braftide was added to HEK293 cell lysate coexpressing V5- and FLAG-tagged BRAF-WT. FLAG-tagged BRAF was pulled down by FLAG antibody-conjugated resin, which was further probed for V5-tagged BRAF. Braftide indeed reduced homodimer BRAF.
4) Delivery of Braftide into HEK cell for BRAF inhibition. Braftide was tagged with cell-penetrating peptide TAT. TAT-Braftide (and its negative control TAT alone) was used to treat HEK293 cells transiently transfected with BRAF-WT and BRAF-G469A. Four hours of treatment resulted in reductions of BRAF, pMEK, MEK (i.e. the MAPK pathway), which were analyzed with respective antibodies by immunoblotting.
5) Delivery of Braftide into cancer cells for BRAF inhibition and cell proliferation inhibition. Two colon cancer cell lines (KRAS-G13D-colon carcinoma) were treated with cell-penetrating TAT-Braftide and assayed for the inhibition of BRAF activity, down-regulation of MAPK signaling, and cell proliferation. All were shown positive, while the negative control TAT alone were negative.
To provide the best experiences, we use technologies like cookies to store and/or access device information. Consenting to these technologies will allow us to process data such as browsing behavior or unique IDs on this site. Not consenting or withdrawing consent, may adversely affect certain features and functions.
Functional
Always active
The technical storage or access is strictly necessary for the legitimate purpose of enabling the use of a specific service explicitly requested by the subscriber or user, or for the sole purpose of carrying out the transmission of a communication over an electronic communications network.
Preferences
The technical storage or access is necessary for the legitimate purpose of storing preferences that are not requested by the subscriber or user.
Statistics
The technical storage or access that is used exclusively for statistical purposes.The technical storage or access that is used exclusively for anonymous statistical purposes. Without a subpoena, voluntary compliance on the part of your Internet Service Provider, or additional records from a third party, information stored or retrieved for this purpose alone cannot usually be used to identify you.
Marketing
The technical storage or access is required to create user profiles to send advertising, or to track the user on a website or across several websites for similar marketing purposes.